| |
| Home | Research | Publications | People | Positions | Multimedia |
|
Research synopsis
We take for granted our ability to perceive the external world, but how does the nervous system accomplish this remarkable feat? Perception begins with physical stimuli that activate sensory cells in the eye, ear, or skin, but it ends with an inference made by the brain. Our sensory organs encode physical stimuli as patterns of electrical impulses that enter the central nervous system, where our neural machinery mysteriously works "behind the scenes" to decode those impulse patterns in order to generate perceptual conclusions. How does this happen? Our laboratory investigates these questions with a focus on the sense of touch. We investigate the full gamut of discriminative touch, which begins with physical stimulation (spatially and/or temporally varying forces on the skin) and ends with perception (the brain's interpretation of the stimulus). Using a combination of experimental and theoretical approaches, we consider perception from the points of view of physics (the mechanical stimulus to the skin), neurophysiology (the neural response), and probability calculus (the perceptual inference). Our research methods range from cutaneous measurement to psychophysical testing to Bayesian computational modeling. We enjoy studying perception in its myriad forms. We attempt to understand fascinating phenomena such as perceptual learning, sensory compensation in blindness, sex differences in tactile acuity, the sense of touch during development and aging, and sensory illusions. An ultimate goal of our research is to formulate mathematically accurate models of human perception that have the predictive power and elegance of the equations of physics. |
|
Innovative psychophysics procedures and device
development
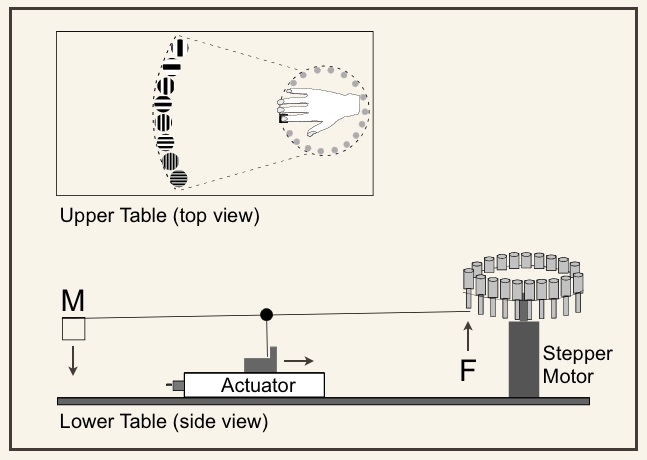
In our psychophysics experiments, we strive to probe tactile perception with precisely controlled stimuli. This has led us to design and build novel equipment when commercial equipment does not exist for a specific purpose. For example, we have invented:
In addition to creating devices for tactile psychophysics research, we evaluate tactile devices for practical use. For instance, we have evaluated a vibrotactile navigational aid for blind individuals (Bharadwaj et al., 2019). We also create and evaluate different tactile testing procedures, with the goal of developing more rigorous and efficient perceptual assessments. For instance, in Tong et al. (2013), we show that an unconventional two-point acuity test, two-point orientation discrimination (2POD), provides a rigorous measure of tactile spatial acuity; we propose that 2POD replace traditional two-point testing in neurological assessment. See our publications: Bharadwaj A, Shaw SB, Goldreich D
(2019) Comparing
Tactile to Auditory Guidance for Blind Individuals. Frontiers
in Human Neuroscience 13:443. doi:
10.3389/fnhum.2019.00443.
Tong J, Mao O, Goldreich D (2013) Two-point
orientation
discrimination versus the traditional two-point test for tactile
spatial acuity assessment. Frontiers in Human
Neuroscience 7:579. doi: 10.3389/fnhum.2013.00579.
Goldreich D, Wong M, Peters RM, Kanics IM (2009) A tactile automated passive-finger stimulator (TAPS). Journal of Visualized Experiments. 28: e1374. doi: 10.3791/1374. |
|
The skin's role in tactile perception
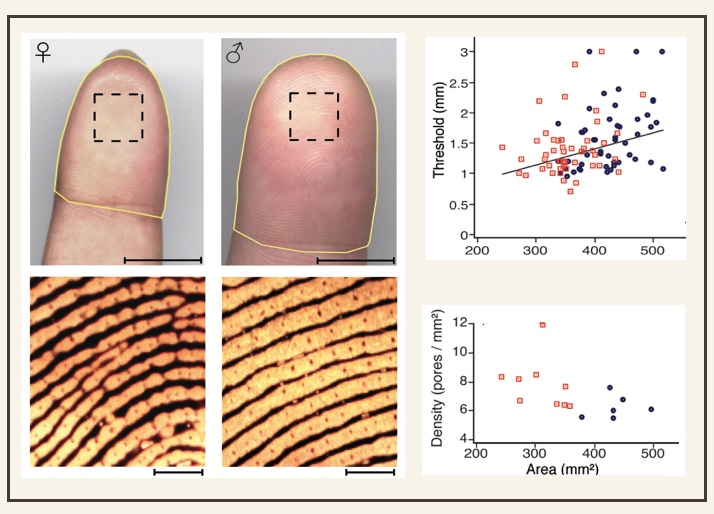 The skin, the body's largest sensory surface, is endowed with a rich array of receptors that detect static pressure, vibration, stretch, temperature, and other stimulus features. We are interested in how the skin and its cutaneous receptors establish the limits of tactile acuity. For example, do people with more compliant skin have better tactile acuity? How does variation in receptor density between individuals affect performance on tactile spatial tasks? To investigate, we measure tactile acuity and skin features in the same individuals. Two skin features of interest are sweat pore density and fingertip surface area. Merkel cells, cutaneous receptors that respond to static skin indentation, cluster in the deep epidermis beneath sweat pores on the skin's surface. We have shown that index fingertip surface area predicts both sweat pore density and tactile spatial acuity, such that people with smaller index fingers have more sweat pores per square mm and finer spatial acuity. We infer that cutaneous receptors are packed more densely in smaller fingers, conferring finer spatial perception. Thus women, for instance, tend to have better tactile acuity than men. The figure above, adapted from Peters, Hackeman, and Goldreich (2009), shows high-resolution images of the index fingertips of a woman and man. Notice that sweat pores (punctate stain) are more closely spaced in the woman's finger. As fingertip surface area increases, sweat pore density decreases (lower right scatterplot) and tactile performance worsens (upper plot). Red symbols: women; blue symbols: men. See our publications: Peters RM, Goldreich D (2013) Tactile
spatial
acuity in childhood: effects of age and fingertip size. PLOS
ONE 8(12): e84650.
Wong M, Peters RM, Goldreich D (2013) A physical constraint on perceptual learning: Tactile spatial acuity improves with training to a limit set by finger size. Journal of Neuroscience 33(22): 9345-9352. Peters R, Hackeman E, Goldreich D (2009) Diminutive digits discern delicate details: fingertip size and the sex difference in tactile spatial acuity. Journal of Neuroscience 29(50): 15756 -15761. |
|
Perceptual learning, plasticity, and development
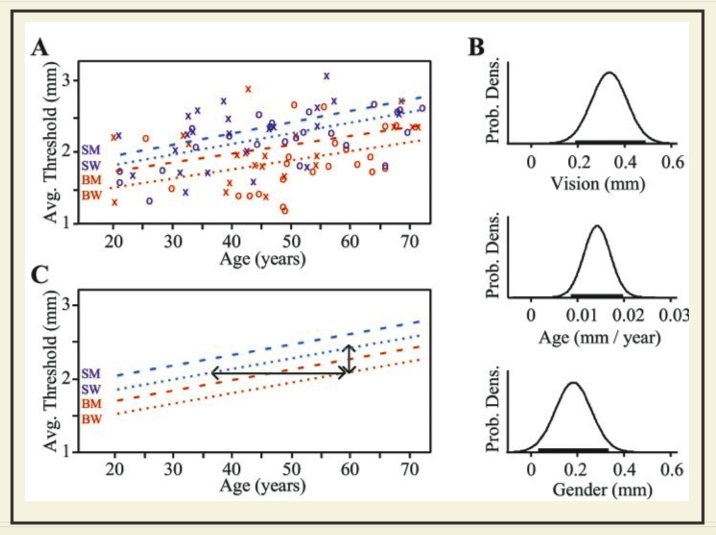 Tactile sensation (the sensorineural response to physical stimuli) begins with the receptors in the skin, but tactile perception (the interpretation of this sensorineural activity) is achieved by the central nervous system (CNS). We are interested in characterizing this central processing of somatosensory information, and understanding how it changes with plasticity, learning, and development. We study how tactile perception changes upon functional reorganization of the central nervous system. For instance, in blind people, normally visual cortical areas acquire tactile responsiveness. What, if any, is the perceptual consequence of this crossmodal plasticity? When blind people learn to read Braille, the representation of the reading finger expands within the somatosensory homunculus. What, if any, is the perceptual consequence of this somatosensory plasticity? The goal of this line of research is to determine the changes in perception that may result from known changes in functional brain organization. The lab's experimental projects in this area include psychophysical studies of tactile spatial acuity and vibrotactile acuity of blind and sighted people. The figure above, from Goldreich and Kanics (2003), shows the effects on index finger tactile spatial acuity of blindness, age, and sex. We study how tactile perception changes with training (perceptual learning), with maturation of the central nervous system (childhood development), and with aging. Some of our studies in these areas have revealed the combined effects of central (training or development) and peripheral (finger size) factors, and are therefore listed also in the section above (the skin). See our publications: Peters RM, Goldreich D (2013) Tactile
spatial
acuity in childhood: effects of age and fingertip size. PLOS
ONE 8(12): e84650.
Wong M, Peters RM, Goldreich D (2013) A physical constraint on perceptual learning: Tactile spatial acuity improves with training to a limit set by finger size. Journal of Neuroscience 33(22): 9345-9352. Wong M, Hackeman E, Hurd C, Goldreich D (2011) Short-term visual deprivation does not enhance passive tactile spatial acuity. PLOS ONE 6(9): e25277. Wong M, Gnanakumaran V, Goldreich D (2011) Tactile spatial acuity enhancement in blindness: evidence for experience-dependent mechanisms. Journal of Neuroscience 31(19): 7028-7037. Bhattacharjee A, Ye AJ, Lisak JA, Vargas MG, Goldreich D (2010) Vibrotactile masking experiments reveal accelerated somatosensory processing in congenitally blind Braille readers. Journal of Neuroscience 30(43): 14288-14298. Goldreich D, Kanics IM (2006) Performance of blind and sighted humans on a tactile grating detection task. Perception & Psychophysics 68(8): 1363-1371. Goldreich D, Kanics IM (2003) Tactile acuity is enhanced in blindness. Journal of Neuroscience 23(8): 3439-3445. |
|
Perceptual inference and illusions
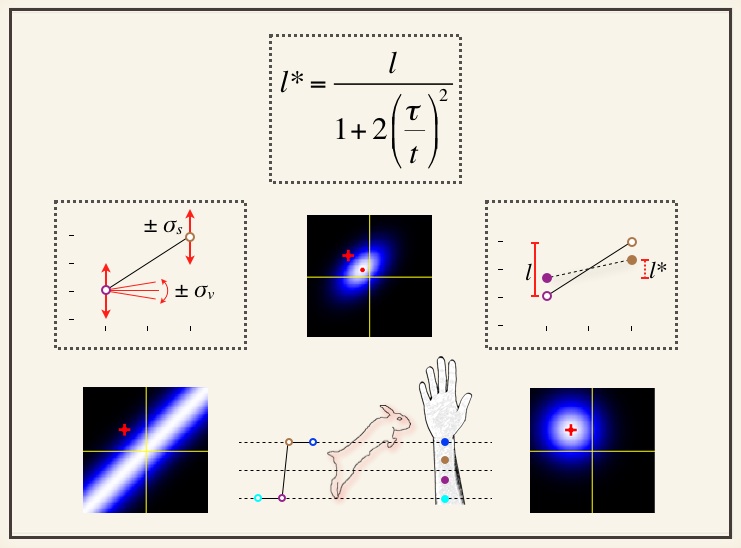 We study tactile perception as a process of Bayesian inference. Our objectives are to create and experimentally test: 1) Bayesian perceptual models of human tactile illusions, and 2) Bayesian ideal observer models that optimally process simulated sensorineural signals, providing a benchmark against which to assess human performance on tactile tasks. In addition, in collaboration with researchers in non-tactile modalities (e.g., vision, audition), we aim to develop and test Bayesian perceptual models relevant to those modalities. The figure above depicts our Bayesian perceptual model that replicates a variety of tactile spatiotemporal illusions, including the tau effect and the famous sensory saltation phenomenon known as the cutaneous rabbit illusion. Our work on this topic is described in Goldreich (2007), Goldreich and Tong (2013), and Tong, Ngo and Goldreich (2016). The model's tactile length contraction formula (top center) relates the perceived distance between two taps to the skin, l*, to the actual distance, l, and time delay, t, between the taps. See our publications: Ma WJ, Kording KP, Goldreich D
(2023) Bayesian
Models of Perception and Action: An Introduction. MIT
Press (ISBN: 9780262047593).
Li L, Chan A, Iqbal SM, Goldreich D
(2017) An
adaptation-induced
repulsion illusion in tactile spatial perception. Frontiers
in Human Neuroscience 11: 331. doi:
10.3389/fnhum.2017.00331.
Tong J, Ngo V, Goldreich D (2016) Tactile
length contraction as Bayesian inference. Journal of
Neurophysiology 116: 369-379.
Peters RM, Staibano P, Goldreich D
(2015) Tactile
orientation
perception: an ideal observer analysis of human psychophysical
performance in relation to macaque area 3b receptive fields.
Journal of Neurophysiology 114: 3076-3096.
Goldreich D, Tong J (2013) Prediction, postdiction, and perceptual length contraction: a Bayesian low-speed prior captures the cutaneous rabbit and related illusions. Frontiers in Psychology (Consciousness Research) 4:221 doi: 10.3389/fpsyg.2013.00221. See also this paper's companion computer program, Leaping Lagomorphs. Goldreich D, Peterson MA (2012) A
Bayesian observer replicates convexity context effects in
figure-ground perception. Seeing and Perceiving (now
Multisensory Research) 25(3-4): 365-395.
Goldreich D (2007) A
Bayesian perceptual model replicates the cutaneous rabbit and
other tactile spatiotemporal illusions. PLOS
ONE 2(3): e333.
|
